Stem rust
COMMON NAME:Stem rust, black rust
SCIENTIFIC NAME: Puccinia graminis Pers.:Pers. f. sp. tritici Eriks. E. Henn.
SYMPTOMS: Uredinia generally appear as oval lesions on leaf sheaths, true stem, and spike. Uredinia can appear on the leaves if other diseases have not killed them. Uredinia are brick red in color and can be seen to rupture the host epidermis, on the leaves uredinia generally penetrate to sporulate on both surfaces. Infected areas are rough to the touch.
ENVIRONMENTAL CONDITIONS: Stem rust is favored by hot days 25-30 C, mild nights 15-20 C with adequate moisture for night time dews. Wind can effectively disperse urediniospores over great distances. Rain is necessary for effective deposition of uredinospore involved in regional spore transport.
INOCULUM SOURCE AND INFECTION:


A B
A. Uredinia of Puccinia graminis f. sp. tritici B. Scanning electron micrograph (SEM) view of a single uredinium


Urediniospores of Puccinia graminis A. normal B. SEM figure


Telia on wheat plants and Teliospores of Puccinia graminis f. sp. tritici. Note dark color and thick cell walls.
On barberry and other alternate hosts:





Use of earlier-maturing wheat varieties in the central Great Plains of the U.S. has helped reduce the threat of stem rust epidemics. Modern wheat varieties in that region mature about 2 weeks earlier than older varieties. This limits the length of time for stem rust epidemics to develop in the central Great Plains as well as the numbers of urediniospores that can contribute to epidemics farther north.
Genetic resistance- Genetic resistance is the most commonly used and the most effective means to control stem rust. Its success is directly linked to the reduced number of races present in the fungal population following the barberry eradication program. Because funding for the program has been reduced in recent years, scientists fear that the remaining barberry bushes will continue to spread into wheat-growing areas to serve both as a source of inoculum and as a means by which the fungus can complete its sexual cycle. The currently used resistance genes should not be expected to remain effective as new races of the fungus begin to appear.
Even without the presence of alternate hosts, the fungus is capable of overcoming resistance genes, primarily through mutation. For this reason, plant pathologists monitor the race populations each year and advise wheat breeders about which resistance genes will best protect the wheat crop in various areas. Wheat breeders use a combination of vertical resistance genes against specific races of P. graminis and horizontal resistance genes that slow the development of the epidemic by offering some resistance to all pathogen races.
Chemical control- In some areas where disease pressure is high, fungicides are applied to wheat to control rust diseases. Fungicides that inhibit the synthesis of sterols [i.e., sterol biosynthesis inhibitors (SBIs) or demethylation inhibitors (DMIs)] are particularly effective, but the cost of application is generally prohibitive for routine use in most wheat-growing areas in the U.S.
Potential approaches to management- Urediniospores infect wheat only through stomata. Scientists have studied how germinating urediniospores locate stomata on leaf surfaces. Although several factors are involved, the germ tube is able to detect the guard cells by their physical dimensions relative to the epidermal cells. Once a stoma is found, an appressorium is produced and infection begins. In the future, it may be possible to breed wheat resistant that is resistant to urediniospore infection because it has epidermal patterns that are not recognized by the fungus.
Stem rust was once the most feared disease of cereal crops. It is not as damaging now due to the development of resistant cultivars, but outbreaks may occur when new pathogen races arise against which the existing kinds of resistance are ineffective. Stem rust remains an important threat to wheat and barley and, thus, to the world food supply. Anton deBary first demonstrated the heteroecious life cycle of a rust fungus with Puccinia graminis, the causal agent of stem rust.


COMMON NAME:Stem rust, black rust
SCIENTIFIC NAME: Puccinia graminis Pers.:Pers. f. sp. tritici Eriks. E. Henn.
SYMPTOMS: Uredinia generally appear as oval lesions on leaf sheaths, true stem, and spike. Uredinia can appear on the leaves if other diseases have not killed them. Uredinia are brick red in color and can be seen to rupture the host epidermis, on the leaves uredinia generally penetrate to sporulate on both surfaces. Infected areas are rough to the touch.
ENVIRONMENTAL CONDITIONS: Stem rust is favored by hot days 25-30 C, mild nights 15-20 C with adequate moisture for night time dews. Wind can effectively disperse urediniospores over great distances. Rain is necessary for effective deposition of uredinospore involved in regional spore transport.
INOCULUM SOURCE AND INFECTION:
Aeciospores from Berberis vulgaris are currently rare, but historically it was an important source of inoculum in northern North America and Europe.
Mycelium or uredinia on volunteer wheat, are the most important source of inoculum in tropical and subtropical climates. Windblown urediniospores are usually from earlier maturing wheat from the south in the northern hemisphere, or from the north in the southern hemisphere.

Urediniospores and aeciospore germinate when in contact with free water. Infection by penetration through the stoma. Penetration requires at least a low light intensity. Germination optimum is 18 C, latent period varies from 10 to 15 days in the field with temperatures of 15-30 C.
HOSTS: wheat and barley, common barberry (and some additional Berberis, Mahoberberis, and Mahonia spp.)
SURVIVAL: Stem rust can survive as teliospores during winter when aeciospores are a major source of inoculum. It generally survives as mycelium or uredinia on volunteer wheat during the non-wheat growing season. Uredinospore can be spread by wind into disease-free areas. Sporulating uredinia are active in tropical and some subtropical areas throughout the winter. Occasional dormant mycelium may survive beneath the snow pack in more northern temperate regions.
SURVIVAL: Stem rust can survive as teliospores during winter when aeciospores are a major source of inoculum. It generally survives as mycelium or uredinia on volunteer wheat during the non-wheat growing season. Uredinospore can be spread by wind into disease-free areas. Sporulating uredinia are active in tropical and some subtropical areas throughout the winter. Occasional dormant mycelium may survive beneath the snow pack in more northern temperate regions.
METHOD OF DISSEMINATION:Urediniospores and aeciospores are wind borne. Teliospores remain with the straw.
HOST RANGE:Stem rust is generally confined to Triticum species, although naturally infected plants of Secale cereale, Hordeum vulgaris, H. jubatum, H. pusillum, Elymus junceus occur. Many genera of the tribe Hordeae are infected when artificially inoculated. Other formae specialis of P. graminis attack many cereals and related grasses, and many species are susceptible to more than one formae specialis.

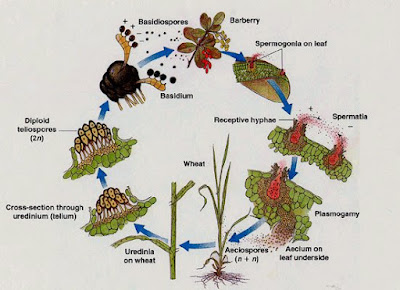
Life cycle
On wheat and other grass hosts:
Plants do not usually show obvious disease symptoms until 7 to 15 days after infection when the oval pustules (uredinia) of powdery, brick-red urediniospores break through the epidermis (Figures). Microscopically, these red spores are covered with fine spines (Figures). The pustules may be abundant and produced on both leaf surfaces and stems of grass hosts. Later in the season, pustules (telia) of black teliospores begin to appear in infected grass species. Microscopically, teliospores are two celled and thick walled (Figure).


A B
A. Uredinia of Puccinia graminis f. sp. tritici B. Scanning electron micrograph (SEM) view of a single uredinium


Urediniospores of Puccinia graminis A. normal B. SEM figure


Telia on wheat plants and Teliospores of Puccinia graminis f. sp. tritici. Note dark color and thick cell walls.
On barberry and other alternate hosts:
Pycnia appear on barberry plants (Figure) in the spring, usually in the upper leaf surfaces. They are often in small clusters and exude pycniospores in a sticky honeydew (Figure). Five to 10 days later, cup-shaped structures filled with orange-yellow, powdery aeciospores break through the lower leaf surface (Figure). The aecial cups are yellow and sometimes elongate to extend up to 5 mm from the leaf surface (Figure). Microscopically, aeciospores have a slightly warty surface (Figure).

Pycnia with honeydew on barberry leaf

Aecial cups on barberry leaf.

Aecial cups and aeciospores on barberry leaf

Aeciospores are produced in chains in the aecium
Disease Management
Barberry eradication:
Barberry eradication:
Once the life cycle of P. graminis was determined, the potential effects of the removal of the barberry alternate host became clear. An expensive and extensive barberry survey and eradication program was initiated in 1918 in the U.S. and continues to a limited extent today.
It was originally hoped that the program would eliminate stem rust as a significant disease in North America, because the basidiospores would have no barberry hosts to infect, and urediniospores could not usually survive harsh winter conditions. The importance of continental spread of stem rust epidemics was not understood until later. Urediniospores overwinter in wheat fields in the southern U.S. and northern Mexico and are then airborne northward via what is now called the "Puccinia Pathway" (Figure). If the weather is favorable for stem rust development in the South, urediniospores will arrive in time and in sufficient numbers to cause epidemics in northern wheat-growing areas.

Each year urediniospores move northward via the "Puccinia Pathway."
Despite this problem, barberry eradication has had significant positive effects on the control of stem rust epidemics. First, it removed a significant, early source of inoculum. A single barberry plant can produce as many as 64 billion aeciospores. Second, it reduced the genetic variation in the fungal population by eliminating the sexual cycle, leaving only asexual urediniospores to maintain the fungus. Mutation is now the primary source of genetic variation. Consequently, there are no longer so many different races of wheat stem rust against which wheat breeders must seek resistance. Finally, epidemics are delayed by several weeks in many of the major wheat producing areas of the U.S. and Canada because aeciospores were released before the first arrival of urediniospores from the south.
Cultural practices- It has long been known that moisture on leaves and excessive foliar nitrogen favor infections by rust fungi. Farmers consider these factors in spacing, row orientation, and fertilizer schedules. Recent changes in production practices may have effects on stem rust. In some areas, summer wheat crops are irrigated, which may increase the survival of infected volunteer plants. In addition, many farmers are practicing no-till or minimum tillage. This increases the probability that rust fungi may successfully overwinter in the protective layer of stubble from the previous crop.
Use of earlier-maturing wheat varieties in the central Great Plains of the U.S. has helped reduce the threat of stem rust epidemics. Modern wheat varieties in that region mature about 2 weeks earlier than older varieties. This limits the length of time for stem rust epidemics to develop in the central Great Plains as well as the numbers of urediniospores that can contribute to epidemics farther north.
Genetic resistance- Genetic resistance is the most commonly used and the most effective means to control stem rust. Its success is directly linked to the reduced number of races present in the fungal population following the barberry eradication program. Because funding for the program has been reduced in recent years, scientists fear that the remaining barberry bushes will continue to spread into wheat-growing areas to serve both as a source of inoculum and as a means by which the fungus can complete its sexual cycle. The currently used resistance genes should not be expected to remain effective as new races of the fungus begin to appear.
Even without the presence of alternate hosts, the fungus is capable of overcoming resistance genes, primarily through mutation. For this reason, plant pathologists monitor the race populations each year and advise wheat breeders about which resistance genes will best protect the wheat crop in various areas. Wheat breeders use a combination of vertical resistance genes against specific races of P. graminis and horizontal resistance genes that slow the development of the epidemic by offering some resistance to all pathogen races.
Chemical control- In some areas where disease pressure is high, fungicides are applied to wheat to control rust diseases. Fungicides that inhibit the synthesis of sterols [i.e., sterol biosynthesis inhibitors (SBIs) or demethylation inhibitors (DMIs)] are particularly effective, but the cost of application is generally prohibitive for routine use in most wheat-growing areas in the U.S.
Potential approaches to management- Urediniospores infect wheat only through stomata. Scientists have studied how germinating urediniospores locate stomata on leaf surfaces. Although several factors are involved, the germ tube is able to detect the guard cells by their physical dimensions relative to the epidermal cells. Once a stoma is found, an appressorium is produced and infection begins. In the future, it may be possible to breed wheat resistant that is resistant to urediniospore infection because it has epidermal patterns that are not recognized by the fungus.
No comments:
Post a Comment